Table of Contents
Sexual reproduction
Most animals reproduce sexually. This means that the genetic material of two individuals (of the same species) is required to produce offspring. Genetically, this is problematic. By combing the genetic material of two parents, the offspring will have more genetic material than its parents. To avoid this, each parent must produce specialised reproductive cells that contain half of the genetic material as the other cells of their bodies. Those reproductive cells are called gametes. The gametes from male parents are called sperm (spermatozoa), while those from the female parent are called eggs (ova). Since the gametes are genetically dissimilar to the other body cells, they are contained in specialised tissues in the testes and ovaries. Hence, they are protected from destruction by the hosts’ immune system (immunologically privileged).
Hormones regulate the production of gametes. In humans, functional gametes are produced after puberty. A male gamete must unite with a female gamete to produce offspring. This union or fusion is called fertilisation. In the animal kingdom, fertilisation can occur within the body of a parent organism (called internal fertilisation) or in the environment (external fertilisation). The following sections will detail these processes.
What happens during fertilisation?
To understand the fertilisation process, we must examine the structure of sperm and egg cells.
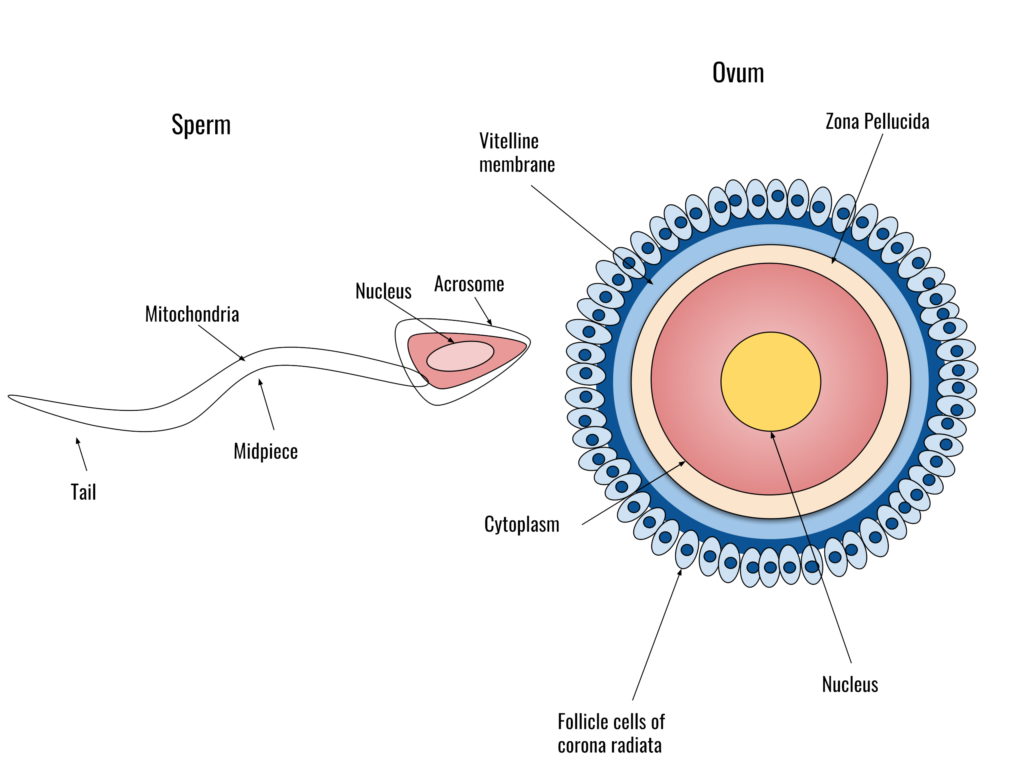
The sperm cell
The human sperm cell is the smallest in the body. It develops from a precursor cell called a spermatocyte. Next, a diploid spermatocyte undergoes meiosis to form haploid spermatids. Finally, the spermatids mature to form sperm cells (spermatozoa).
A sperm cell is shaped like a tadpole and consists of three morphologically-distinct parts: the head, mid-piece and tail:
- Head: The head contains the nucleus, which has a haploid set of chromosomes. It also contains some enzymes needed for fertilisation in the acrosome.
- Midpiece: the midpiece contains mitochondria (50-75 mitochondria per cell). They provide energy for movement.
- Tail: the tail, also called the flagellum, moves in a whip-like manner to propel the cell forward. It contains numerous cytoskeletal proteins.
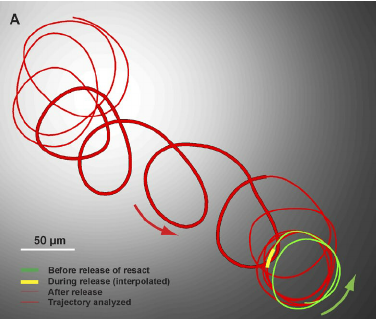
The sperm cell is motile and can move independently. Once released, the cell moves by the beating of the flagella. Sperm cells are chemically attracted to the egg cells – they detect gradients of molecules released from the egg cell’s outer membranes. This type of movement is called chemotaxis. Also, unlike bacterial cells, sperm cells generally move in a circular or spiral trajectory (not a straight trajectory).
The egg cell
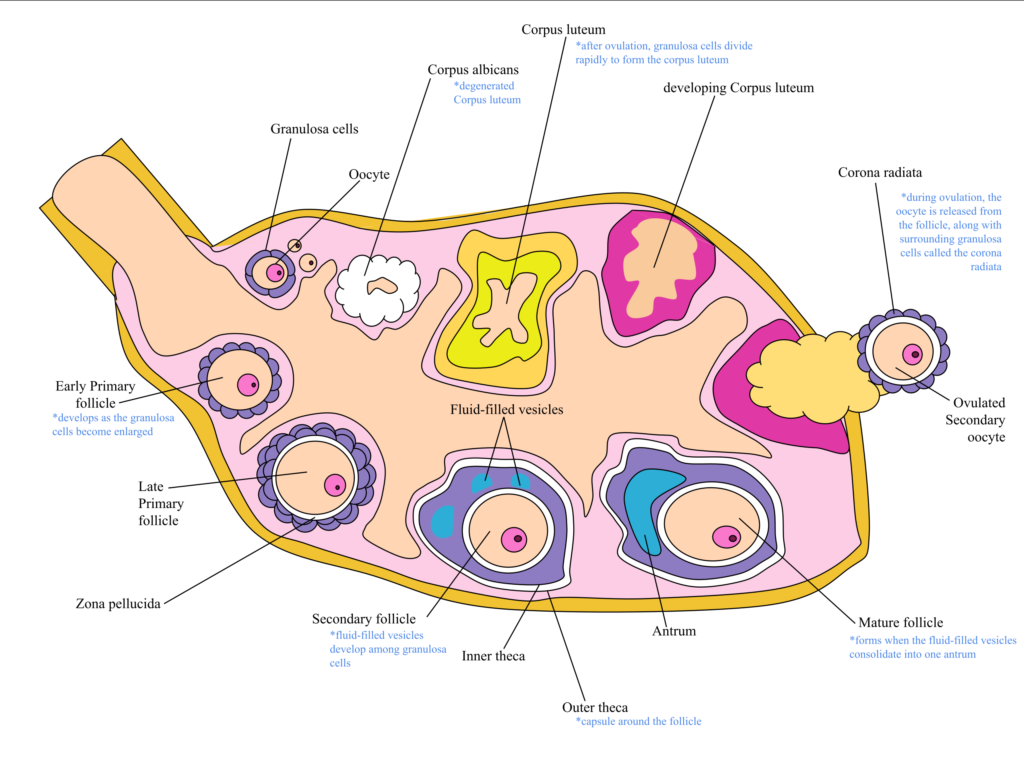
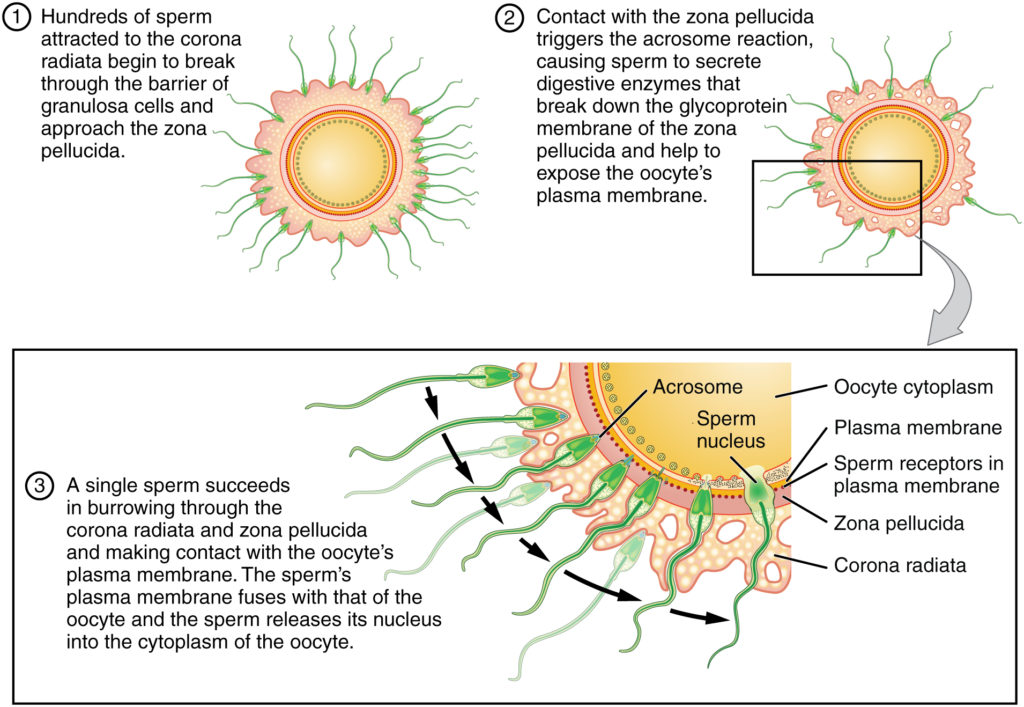
Egg cells are among the largest cells. They are produced in the ovaries of females – diploid oocytes undergo meiotic divisions to produce haploid ova (singular: ovum). However, unlike spermatogenesis, oogenesis is more complex. During early fetal development, cells called oogonia begin to develop into primary oocytes. Each oocyte is surrounded by cells called the granulosa. Together, the oocyte and the surrounding granulosa cells are referred to as a follicle. The early follicle is called the primary follicle. The primary oocyte (i.e., the egg cell within the primary follicle) begins meiosis I but does not complete it (this is referred to as the prophase arrest). Each female baby has, on average, two million primary oocytes in each ovary at birth. Many of these primary oocytes will die off during early development: at the start of puberty, each ovary will contain about 300 000 to 400 000 primary follicles. One primary follicle will be recruited for maturation at each menstrual cycle from puberty onwards, under the influence of Leutinising Hormone (LH, secreted by the anterior pituitary gland). During maturation, the primary oocyte will complete meiosis I and begin meiosis 2. However, it will not complete meiosis II but stop at metaphase 2 (metaphase arrest). At this stage, the follicle is referred to as a secondary follicle. The oocyte then becomes detached from the rest of the follicle and is surrounded by a group of cells called the corona radiata. Under the influence of the LH hormone (LH surge), the secondary follicle will rupture, releasing the ovum (egg) into the Fallopian tube. This process is ovulation. The remains of the follicle become another structure in the ovary – the corpus luteum. The corpus luteum becomes an endocrine tissue and secretes the hormone progesterone, preparing the endometrium for implantation.
Fertilisation
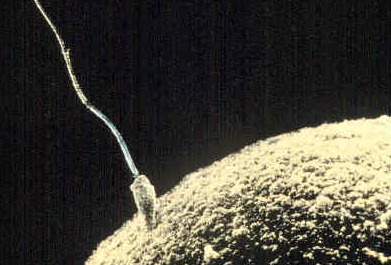
Fertilisation is the union of gametes to produce a zygote. The basic processes of fertilisation are common to sexually-reproducing animals. As indicated above, sperm cells move chemotactically towards egg cells. When they encounter the egg cells, they will initially attach loosely to the corona radiata (these are cells surrounding the oocyte). Next, the sperm cells will push through this layer and adhere to the zona pellucida. The zona pellucida contains various transmembrane proteins, collectively referred to as ZP proteins. Proteins on the sperm cell surface bind to the ZP proteins (specifically the ZP3 protein). ZP3 is highly species-specific – this is why it is not possible for gametes from different species to fertilise each other). The attachment to the ZP3 protein triggers the acrosome reaction in the sperm cell: this causes hydrolytic enzymes to be released from the sperm head. The enzymes digest the zona pellucida layer in the vicinity of the attached sperm. Once the zona pellucida has been breached, the sperm cell membrane fuses with the egg cell’s plasma membrane. This causes the sperm cell’s nucleus to be released into the egg cell’s cytoplasm. After that, the sperm nucleus will fuse with the egg nucleus, thus creating a diploid cell. This completes fertilisation, and the resultant cell is called a zygote.
Of the many sperm cells that initially attach to the corona radiata, only one will successfully attach to a ZP3 protein and initiate the acrosome reaction. The acrosome reaction also initiates a cortical reaction in the egg, where the ovum retracts from the zona pellucida. This prevents other sperm cells from fertilising it (i.e. preventing polyspermy).
Further thoughts on fertilisation
In many species, male and female gametes differ widely in shape and structure (as discussed above). Furthermore, they differ in their sizes, and, generally, females produce fewer gametes than males. The differences between male and female gametes are referred to as anisogamy. Scientists think anisogamy evolved because males and females faced different selective pressures to increase their reproductive success. Males seek to maximise their number of offspring by producing small gametes in large quantities. However, females produce fewer, larger gametes. This is because the egg carries all the nutrients and developmental factors that the embryo needs during the early stages of life. Both strategies are directed at increasing the chances of survival of the progeny.
Because of genetic changes that occur during meiosis, not all sperm cells from a male organism are genetically identical. Since only one of many millions of sperm cells will fertilise an egg cell and pass on its genetic legacy to the progeny, scientists believe that sperm cells compete with each other. This is referred to as sperm competition. Sperm competition may have evolved due to non-monogamous mating (where a female may be impregnated by multiple males – polygyny and polygynandry). Sperm competition is a form of sexual selection, a potent driver of evolutionary change in species.
Locating fertilisation
Now that we have explored fertilisation a little, we will look at where fertilisation occurs in sexually-reproducing organisms. Biologically, fertilisation is internal if it occurs within an individual’s body. Otherwise, fertilisation is external. Both refer to fertilisation in the natural setting: if fertilisation is conducted in a laboratory, it is called in vitro fertilisation.
Internal fertilisation
In internal fertilisation, gametes are released into the reproductive tissues of the female parent. Therefore, the female parent possesses reproductive structures that produce eggs (ovaries) and allow for the transfer of the eggs to the sites of fertilisation (oviduct or Fallopian tube). If the embryo develops internally within the parent, then there will be reproductive tissues that support it (uterus). The male parent contains reproductive tissues that produce sperm (testis) and various structure for delivering the sperm into a female mate’s reproductive system.
In species that perform internal fertilisation, there are variations in the way the embryos develop:
- Oviparity: After fertilisation, females lay the eggs containing the undeveloped embryo. The embryo develops externally within the egg and will hatch once it attains developmental maturity.
- Viviparity: In viviparity, the embryo develops within the parent. Live young progeny are delivered at birth.
- Ovoviviparity: In this mode of reproduction, embryos develop within eggs, but those eggs remain within the parent until birth.
Advantages of internal fertilisation
- Mate choice: females generally are able to choose the best mate for copulation and production of progeny. In species where internal fertilisation is the norm, elaborate mating rituals are common.
- Protection: both gametes and the developing embryos are protected from environmental factors, such as heat and water availability (dehydration). Also, in many species, the young progeny are protected from predation (until they have attained developmental maturity).
Disadvantages of internal fertilisation
- Carrying the undeveloped embryo within her body can be dangerous for the female parent. This is because she must provide nutrients to the developing embryo (energy costs), and the growing embryo (during gestation) can cause physiological and other forms of damage to her.
- At each reproductive cycle, only a small number of offspring are produced. This can significantly impact population sizes when conditions fluctuate.
External fertilisation
In this mode of reproduction, male and female parents release their gametes into the environment where fertilisation occurs. Once the gametes are released, the parents have little control over how and if fertilisation will occur. Fertilisation success is dependent on favourable environmental factors that allow gametes to come into contact with each other. Therefore, large numbers of gametes are produced in species that employ external fertilisation.
Broadcast spawning
Sessile aquatic invertebrates largely carry out broadcast spawning. The males and females of these species release their gametes into the water column. Water currents transport the gametes some distance away from the parents. Fertilisation occurs in the water column, and after early development, the embryos settle on some surface to continue their development into their juvenile and adult forms. For this method of reproduction to be successful, male and female animals must release their gametes at the same time. Various environmental variables may act as triggers. For example, some organisms spawn seasonally, while others may respond to changes in temperature, pH or chemicals in the environment.

Motile aquatic organisms engage in mating behaviours to enhance fertilisation success. For example, in amphibians, mating ensures that the males release their sperm directly onto the eggs as soon as the latter are laid in the water. Other species exhibit brooding behaviours to maximise the survival of the progeny after external fertilisation.
Advantages of external fertilisation
- Species that reproduce through external fertilisation usually produce a large number of gametes and hence, potentially a large number of offspring. However, many of those progeny will not survive to adulthood.
- The mixing of the gene pool for externally fertilising species may occur to a greater extent than that for internally fertilising species. This is because of the large numbers of gametes released into the environment during breeding.
- Because they use various abiotic and biotic media for gamete dispersal, these species may be better colonisers (that species that adopt internal fertilisation) and occupy larger habitats.
- The female parent does not have to face the dangers of gestation.
Disadvantages of external fertilisation
- Reproductive success is highly dependent on environmental and other factors (i.e. biotic and abiotic). Generally, fertilisation rates are low.
- As early development occurs externally (relative to the parent organism), the chances of the immature young making adulthood are low.