Inquiry question: How important is it for genetic material to be replicated exactly?
Students model the processes involved in cell replication, including but not limited to mitosis and meiosis
Introduction
One property of living things is their ability to reproduce. In the previous inquiry question, we explored the different strategies various organisms use to give rise to the next generation. For example, single-celled organisms divide to produce single-celled progeny. Similarly, the progeny of multicellular organisms is also multicellular. However, even multicellular organisms commence life as single cells.
During reproduction, the genetic material is passed on to the progeny. This means that the genetic material in the parental organisms must be copied before it is transferred to the progeny cells. But how faithful is the replicated DNA to the original? The answer depends on the type of reproduction that occurs:
Asexual reproduction: For cellular divisions such as binary fission and mitosis, the original and copied DNA is nearly identical
Sexual reproduction: For meiosis, the replicated DNA is a slightly altered version of the original.
The Cell Cycle
Knowing what happens to cells when not dividing is essential to understanding cell division. The lifetime of a single cell is referred to as its cell cycle. The cell cycle is "a series of events that occurs in a cell as it grows and divides" (NHGRI). The cell cycle is divided into three non-dividing phases, collectively called interphase, and one dividing phase (mitosis or meiosis). The three non-dividing phases of interphase are Synthesis (S) and two rest phases called Gap (G1 and G2). The cell division phase is abbreviated M. This is shown in the following figure.
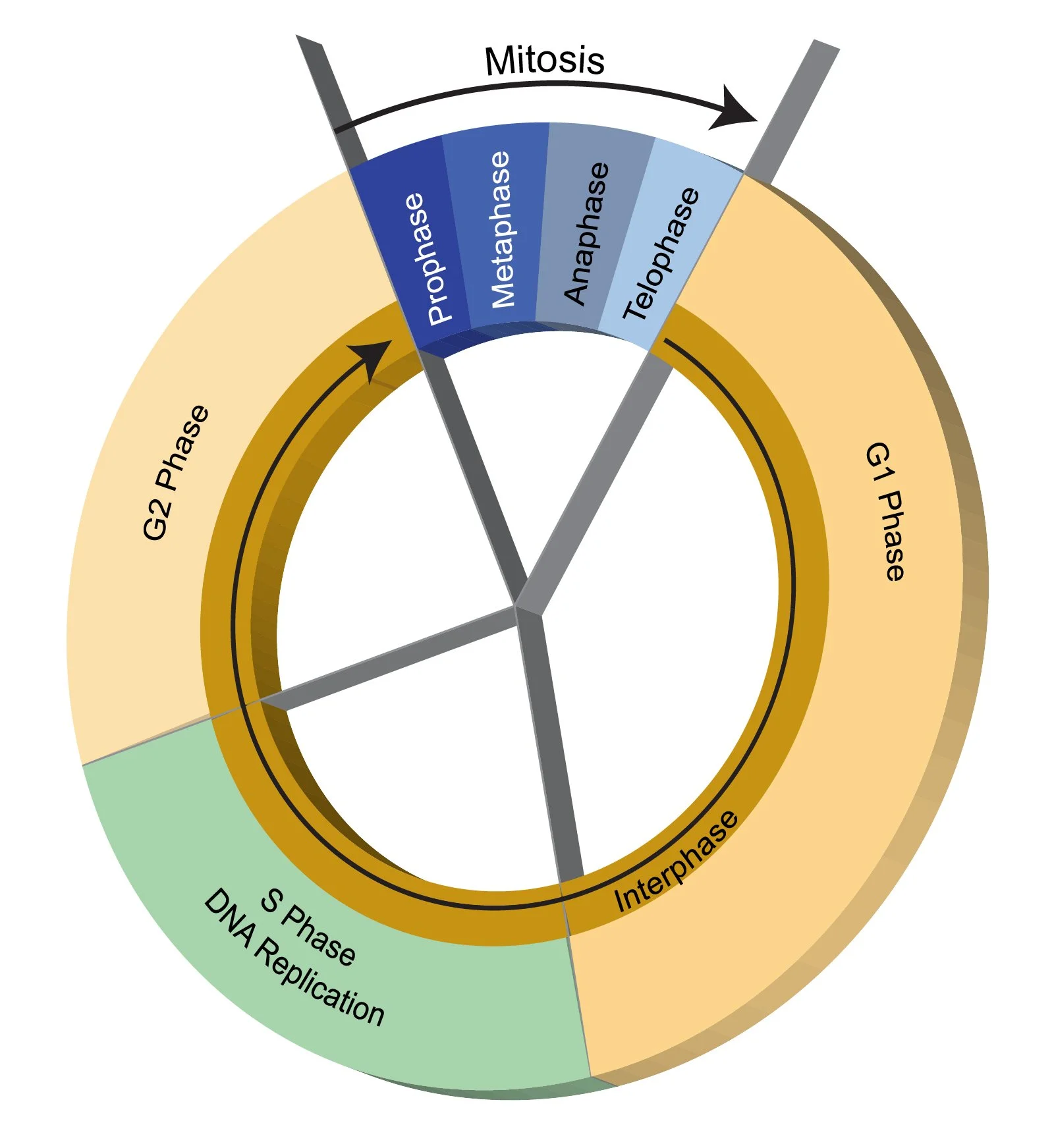
The stages of the cell cycle. After a cell completes cell division (M), each daughter cell enters the G1 phase. During G1, the cells grow in size. Then, during the S phase, each cell duplicates its genome. During the G2 phase, the cells prepare for cell division. When ready, the cells enter the M phase, where they divide. The cell cycle is unidirectional - cells cannot move backwards in the cycle. Image credit: NGHRI
Mitosis
Cells enter mitosis after completing G2. Mitosis is the beginning of cell division, which has two distinct parts: nuclear and cytoplasmic. Nuclear division (mitosis) occurs before cytoplasmic division (cytokinesis). Based on the behaviours of the chromosomes, mitosis consists of four distinct phases: prophase, metaphase, anaphase and telophase.
Before the cell enters mitosis, it duplicates its entire genome. For example, a human cell's genome consists of 46 chromosomes arranged in 23 pairs. However, when the cell enters mitosis, it has 92 chromosomes (46 x 2), arranged as 46 bivalent chromosomes. The bivalent chromosomes are not visible until the cell is in late prophase.
Prophase
Prophase is the first stage of nuclear replication. The main cellular events of prophase are;
Condensation of chromosomes: chromosomes become tightly packaged around histone proteins. Condensed chromosomes become visible under the microscope. In addition, condensed chromosomes take up some nuclear stains and appear like short pieces of thread (the word 'mitosis' means thread in Greek).
Duplication of centrioles: Centrioles are protein structures that form long protein strands called spindle fibres. They move to opposite sides of the cell and define the positions of the replicated nuclei. Also, during cytokinesis, the cytoplasm divides in the region between the centrioles.
Spindle fibres: the spindle fibres elongate from the centrioles. They will eventually attach to the chromosomes. This process is so precise that one fibre from each centriole will attach to one chromatid of a bivalent chromosome.
Breakdown of nuclear envelope: The nuclear envelope is made up of two membranes—that is, it is double-membraned. The membrane disintegrates and forms tiny circular fragments that attach to the chromosomes.

A figure of a bivalent chromosome. It consists of two chromatids (also called sister chromatids), that are joined together at the centromere. Each chromatid is a single molecule of DNA. The sister chromatids of a bivalent chromosome are identical (in terms of their DNA sequences). Image credit: Genome.gov
By the end of prophase, condensed bivalent chromosomes are seen to be attached to spindle fibres. The growing spindle fibres push the chromosome towards the centre of the cell. This signifies the end of prophase and the start of metaphase.

The cell in the middle of this image is in late prophase. The chromosomes are condensed and visible as thread-like structures. The nuclear membrane has broken down and is not visible. The centrioles and spindle fibres are also not visible. Image credit: This image comes from Josef Reischig's archive and is one of the 384 pictures kindly donated by the authorship heirs under a CC BY SA 3.0 license as part of Wikimedia Czech Republic's GLAM initiative.

An overview of chromosome condensation. Initially, DNA is wrapped around histone protein complexes (1). The histone-DNA units (nucleosomes) are then tightly packed to form the condensed chromosome (2-4). Another protein complex, called cohesin, holds the sister chromatids together. Image credit: Tatsuya Hirano (RIKEN). ©RIKEN

During DNA replication, proteins called cohesins (green circles) attach to the DNA molecules (1). After replication, the cohesins attach the daughter DNA molecules together (2). Each daughter DNA molecule is a sister chromatid. During mitosis, condensin protein complexes (red circles) bind to the DNA molecules, thus initiating DNA condensation (3). At anaphase, the cohesin proteins fall off the DNA molecules. This causes the sister chromatids to separate and be pulled to opposite ends of the cell by the spindle fibres (3-4). Image credit: Tatsuya Hirano (RIKEN). ©RIKEN

Meters of DNA are packed into micrometre-sized chromosomes in mitosis, shown here in mitosis-arrested human (HeLa) cells. Chromosomes were spread and stained with a DNA-binding dye. Scale bar, 10 μm. Uhlmann, F. Open questions: Chromosome condensation - Why does a chromosome look like a chromosome? BMC Biol, 11, 9 (2013). https://doi.org/10.1186/1741-7007-11-9

Chromosomes of dinoflagellates. The nuclei are stained with Hoechst 33258 (blue) (A) Karenia brevis, (B) Karenia mikimotoi, (C) Karlodinium veneficum, and (D–I) Karenia selliformis nuclei at different stages of interphase (D–F) or during division (G–H). (I) A set of chromosomes at 2× magnification. By individually following some of the chromosomes, their bands can be counted. Scale bar = 10 μm. Image credit: Figueroa RI, de Bustos A, Cuadrado Á (2018) A novel FISH technique for labelling the chromosomes of dinoflagellates in suspension. PLoS ONE 13(10): e0204382. https://doi.org/10.1371/journal.pone.0204382. CC 4.0.

Fluorescence micrograph of a cell in early prophase. The condensed chromosomes are shown in blue, while the proteins that make up the spindle fibres are green. Note that the spindle fibres elongate from the centrosome and will attach to the centromeres of chromosomes. Fragments of the nuclear envelope will attach to the chromosomes. Image credit: © Michael Davidson. Observing Mitosis with Fluorescence Microscopy.

The figure shows a bivalent chromosome attached to spindle fibres at the centromere. The kinetochore is a protein complex at the centromere, and the attachment to microtubules occurs here. Image credit: One Ring to Bind Them All Nature's Smart, Elegant Solution to Preserving Genetic Integrity. Science@Berkeley Lab.

A model of nuclear envelope disassembly. The nuclear envelope is shown in grey, while the nuclear pores are shown as blue discs. During prophase, the centrosomes form and produce spindle fibres (microtubules), some of which attach to the nuclear envelope. As the centrosomes separate, the movement of the spindle fibres 'tears' the nuclear envelope. This exposes the nuclear lamina (the cytoskeletal system inside the nucleus. The lamina is shown as the purple structures). As the envelope tears, the lamina breaks down. This process continues until the entire nuclear structure disintegrates. The condensing chromosomes (orange) attach to spindle fibres. Image credit: Aitchison JD, Rout MP. A tense time for the nuclear envelope. Cell. 2002 Feb 8;108(3):301-4.
Prometaphase
Some authors define the late prophase and early metaphase as a single distinct mitotic phase, prometaphase. This article does not discuss prometaphase.
Metaphase
In metaphase, the chromosomes are aligned in the middle of the cell. This region is called the metaphase plate and is approximately halfway between the two centrioles. As mentioned in the previous section, the movement of the chromosomes is caused by the lengthening of the spindle fibres.
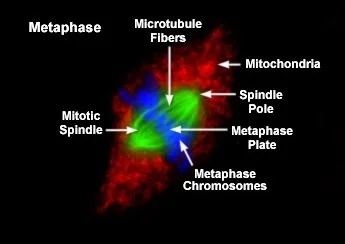
Fluorescence micrograph of a cell in metaphase. The chromosomes (blue) are arrayed in the middle of the cell (metaphase plate). Each bivalent chromosome is attached to a spindle fibre (green) from opposite centrosomes (spindle poles). Image credit: © Michael Davidson. Observing Mitosis with Fluorescence Microscopy
Anaphase
The main event in anaphase is the separation of sister chromatids. The sister chromatids of bivalent chromosomes are held together by proteins that function like glue to hold the replicated DNAs together. At the start of anaphase, specific proteases are activated, resulting in the cleavage of proteins that hold the chromatids together. Then, the spindle fibres contract, pulling the chromatids towards the centrioles. Thus, sister chromatids are separated and move to opposite ends of the cell.

Fluorescence micrograph of a cell in anaphase. Contraction of the spindle fibres separates the chromatids (daughter chromosomes) to opposite spindle poles. Image credit: © Michael Davidson. Observing Mitosis with Fluorescence Microscopy
Telophase
Several things occur during telophase.
Reformation of the nuclear envelope: the membrane fragments associated with the chromosomes fuse. This results in the nuclear membrane forming around the chromosome at the two poles of the cell.
Decondensation of the chromosomes: The chromosomes will unwrap from the histone proteins (not completely). Individual chromosomes are no longer distinctly visible.
Dismantling of centrioles and spindle fibres: These structures, which were responsible for separating sister chromatids, are no longer needed by the cell and are dismantled.
By the end of telophase, a cell will have two distinct nuclei.

Fluorescence micrograph of a cell in telophase. The separation of the daughter chromosomes is complete, and the nucleus reforms. The microtubules and the centrosomes disassemble while the chromosomes decondense. Image credit: © Michael Davidson. Observing Mitosis with Fluorescence Microscopy
Cytokinesis
Cytokinesis is not a mitotic phase but a distinct event after mitosis. It refers to the separation or division of the cytoplasm, which happens in the region between the two nuclei. Contraction of the cytoskeleton in the middle of the cell (near the metaphase plate) causes the plasma membrane to be pulled to the cell's interior, creating a 'cleavage furrow'. In plants, the cytoplasmic division also includes the formation of a cellulose cell wall.

Fluorescence micrograph of a cell in cytokinesis. The separation of the cytoplasm is nearing completion. The nuclei have reformed in both daughter cells. Image credit: © Michael Davidson. Observing Mitosis with Fluorescence Microscopy

Cytokinesis in a dividing sea urchin cell. This scanning electron micrograph shows the cleavage furrow - the inward pinching of the plasma membrane. As a result, the cytoplasm becomes divided between the two newly-formed nuclei, thus forming two distinct cells. Image credit: Spiegel, Evelyn & Howard, Louisa. Center for Research in Biological Systems. The image is in the public domain
Key points
Genomic duplication occurs before mitosis. In the S-phase of the cell cycle, each DNA molecule is replicated. However, the replicated DNA molecules do not separate but remain attached as sister chromatids.
After mitosis, each daughter cell has the same number of chromosomes as its parental cell.
In some organisms, such as some fungal species, cytokinesis does not occur after mitosis. Instead, the two nuclei in the cell will undergo further rounds of mitosis, resulting in large, multi-nucleated cells.

Image of mitosis in sea urchin cells. Chromosomes are shown in blue, while proteins (spindle fibres are in gold).Image credit: von Dassow, Bement, Foe, Odell. Cytoskeletal Dynamics During Cytokinesis. Center for Cell Dynamics
Changes in DNA content during the cell cycle
The amount of DNA a cell changes during the cell cycle. The cell's 'normal' content of DNA is evident during G1. In the S-phase, the DNA content increases because of DNA replication. By the end of the S phase, the cell's DNA content is double the amount found in the G1 phase. This situation continues until the cell completes G2 and enters mitosis. After mitosis and cytokinesis, the cell enters G1, and its DNA content returns to 'normal'.

A graph showing the changes in the DNA content of a cell during the cell cycle. The cell's 'normal' DNA content is arbitrarily denoted as 1. DNA replication during the S phase increases the cell's DNA content. The cell's DNA content returns to normal after mitosis (M phase). Image credit: Darzynkiewicz, Zbigniew & Halicka, H & Zhao, Hong. (2010). Analysis of Cellular DNA Content by Flow and Laser Scanning Cytometry. Advances in experimental medicine and biology. 676. 137-47. 10.1007/978-1-4419-6199-0_9.
The role of mitosis
In unicellular eukaryotic organisms, mitosis increases population size. Thus, mitosis functions as organismic (whole-organism) reproduction in those instances. In multicellular eukaryotes, mitosis contributes to:
Increase in organism size - this is due to the increase in cell numbers in multicellular organisms
Tissue repair and regeneration: Mitosis replaces cells that die (apoptosis and necrosis) or are damaged. This often occurs as a result of mitotic divisions of stem cells.